Rafael Garcia publicou um artigo no caderno
Mais! da
Folha de São Paulo, de 06/08/2006 intitulado
"Vestígios de um império" sobre um mecanismo de correção descoberto no RNA que reforçaria seu papel de desbravador da vida no planeta Terra,descrito no estudo de
Nikolay Zenkin, da Universidade Rutgers (EUA) e colegas publicado na revista
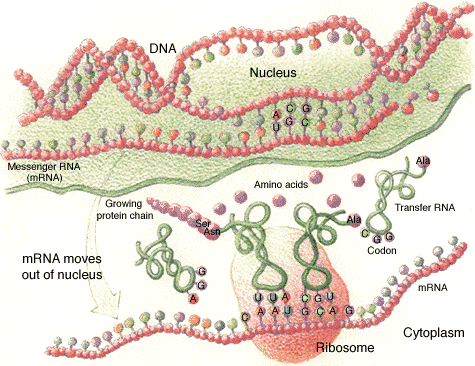
"Science" da semana passada. Ele mostrou em laboratório como a RNA polimerase, a enzima que produz as cópias da molécula, interage com íons de magnésio e moléculas de água num esquema para cortar as partes erradas da do RNA acrescentadas no processo de reprodução.
Infelizmente Rafael Garcia teima em polarizar a questão como sendo
"blablablá criacionista, que com freqüência recorre à falácia lógica do argumento da ignorância – tomar uma premissa como falsa por causa da ausência de provas de que ela é verdadeira".
Rafael,
a questão hoje em dia, e a turma do caderno Ciência da FSP, a Grande Mídia Tupiniquim, mais a Nomenklatura científica, estão cansadas de saber –
não é se as especulações das atuais teorias da origem e evolução da vida contrariam relatos de criação das tradições religiosas, mas se as evidências encontradas na natureza as apóiam ou não. Não apóiam, e estão deixando os naturalistas filosóficos – verdadeiros metafísicos – desesperados, pois não há resposta naturalista que satisfaça o rigor do método científico: Empirica empirice tratanda [As coisas empíricas são empiricamente consideradas]! Aqui nós estamos no 'império do faz de conta'!Ainda bem que Rafael reconhece no seu texto que o trabalho de Zenkin et al.
"não significa que a explicação da origem da vida esteja próxima". Ele está coberto de razão.
Para demonstrar que o trabalho de Zenkin et al ainda está longe de ser uma explicação plausível sobre a origem da vida, chamo a atenção da Nomenklatura tupiniquim para a dificuldade da questão, que não é
'blá blá blá de criacionistas' e nem da turma perversa do Design Inteligente, mas questão científica.
Quem tiver uma teoria da origem da vida mais plausível do que a atual, sugiro submeter seu trabalho à
The Origin of Life Foundation e concorrer a um prêmio de
US$ 1.000.000,00 (Hum milhão de dólares). As exigências epistêmicas são extremamente rigorosas. Há dois professores brasileiros como revisores:
Ricardo Ferreira, Departamento de Quimica Fundamental, UFPE,[meu velho e respeitável oponente] e Romeu Cardoso Guimaraes.
Os leitores me desculpem, mas o texto segue em inglês, pois este blogger está sem tempo, pois retomou sua dissertação e não vai poder publicar freqüentemente neste blog:
The Origin-of-Life Prize ® is offered through The Gene Emergence Project ® of
The Origin-of-Life Foundation, Inc. ®life@us.netDescription and Purpose of the Prize"The Origin-of-Life Prize" ® (hereafter called "the Prize") will be awarded for proposing a highly plausible mechanism for the spontaneous rise of genetic instructions in nature sufficient to give rise to life. To win, the explanation must be consistent with empirical biochemical, kinetic, and thermodynamic concepts as further delineated herein, and be published in a well-respected, peer-reviewed science journal(s).
The one-time Prize will be paid to the winner(s) as a twenty-year annuity in hopes of discouraging theorists' immediate retirement from productive careers. The annuity consists of $50,000.00 (U.S.) per year for twenty consecutive years, totalling one million dollars in payments.
The ability of the Foundation to underwrite these payments and to administer the Project is monitored by the well-known accounting firm of Young, Brophy & Duncan, PC, Certified Public Accountants.
Formal application by submitters is required to win. Submitters must expressly consent to abide by all terms and conditions of the Prize before judging of their paper(s) can begin.
Topic relates to: Rules for The Origin of Life Prize; The Gene Emergence Project; The Origin of Genetic Instructions Prize; Chemical Evolution; Prebiotic Evolution; Abiogenesis; Self-replication; Self-organization; Auto-catalysis; Mutual Catalysis; Abiotic Synthesis; Protobiont; Protocell; Protometabolism; Primordial Milieu; Astrobiology; Exobiology; RNA World; Protein World; Coevolution; Biopoesis; Biogenesis; Lipid world; Homochirality; Bilipid; Self-ordering; Complexity Theory; Artificial Life; Alife; Minimalist Genomes; Minimal Genomes; Code Origin; Genetic Code Theory; Protogene Theory; Minigene; Mini Gene; Clay Matrix; Mineral Adsorption; Bioinformation Theory; Semiotics; Biomessage Theory; Message Theory; SETI; Emergent Properties; Initial Gene Emergence; Biological Information Theory; Aperiodic Specified Complexity; Specified Aperiodicity; Molecular evolution; Messenger Molecules; Information Theory; Chaos Theory; Function Theory; Biofunction Theory; Instruction Theory; Linguistic Theory; Irreducible Complexity; Biopolymers; Thermophiles; GARD; Extremophiles; Hyperthermophiles; Archaea; Prokaryotes; Neural Nets; Decision Theory; Artificial Intelligence; AI; Polarized light from neutron stars.
Currently accepting submissions
Click on the table of contents on the left side of the screen for application requirements, submission forms, criteria for winning,etc.
Other than announcements in scientific journals, The Prize will not be publicly advertised in lay media. The Origin-of-Life Foundation, Inc. wishes to keep the project as quiet as possible within the scientific community. No media interviews will be granted until after the Prize is won.
Purpose of the Prize
"The Origin-of-Life Prize" ® is being offered to stimulate research into chaos, complexity, information, probability, self-organization, and artificial life/intelligence theories as they relate directly to biochemical and molecular biological reality. The Foundation wishes to encourage the pursuit of natural-process explanations and mechanisms of initial "gene" emergence within nature. The subject of interest is the genesis of primordial functional information itself rather than its physico-chemical matrix of retention and transmission. Bioinformation fits into the category of "prescriptive information" ("instruction," rather than mere probabilistic combinatorics [Abel, 2000]). By what mechanisms do stochastic ensembles acquire instructive/integrative potential? In other words, what are the processes whereby random biopolymeric sequences self-organize into indirect, functional code?
Central questions of interest relate to the definition and nature of "genetic instructions" and "biomessage." Is genetic recipe adequately represented and described by "mutual entropy" (shared, correlative uncertainty between transmitter and receiver)? At what point and by what processes do "biofunction" and "biosystem" enter into the chemical evolution of bioinformation?
What is a reasonable, empirically-accountable definition of "minimal life"?
How does nature's genetic programming achieve such long sequences of highly functional decision-node selections?
Genes are linear, digital, quaternary decision-node strings. Nucleotide selections represent four-way algorithmic switch-settings. These switch-settings are covalently-bound into primary structure. The string's specific sequence precedes secondary and tertiary folding. Folding results from forces such as hydrogen bonding, charge attractions/repulsions, and hydrophobicity. These forces are much weaker than the covalent binding that has already determined sequence. Folding space is primarily constrained by this pre-existing nucleotide sequencing. Ultimately, the algorithmic programming instantiated into the nucleotide-selection sequence determines biofunction.
The problem is that natural selection works only at the phenotypic level, not at the genetic level. Neither physicochemical forces nor environmental selection choose the next nucleotide to be added to the biopolymer. Mutations occur at the genetic level. But environmental selection occurs at the folding (functional) level, after-the-fact of already strongly set sequence, and after-the-fact of already established algorithmic function of the folded biopolymer.
By what mechanism did prebiological nature set its initial algorithmic switch-settings to program the first few (RNA?) genes?
How was RNA folding function anticipated when covalently-bound primary structure was forming?
Suppose a self-replicative oligoribonucleotide analog sequence occurred spontaneously out of sequence space. How did this self-replicative strand simultaneously anticipate folding needs for metabolic utility? Any evolution toward folding fitness would tend to mutate the sequencing away from self-replicative fitness. What was the bridge between both functions?
How could random mutations simultaneously contribute to both disparate functions?
How did so many biochemical pathways get integrated into one coherent, unified, and sophisticated metabolic process?
Clarification of what the Foundation is looking for
We are primarily interested in how certain linear digital sequences of monomers acquired three-dimensional dynamic function. The Prize offer is designed to stimulate focused research on the origin of initial genetic instructions themselves. So much of life origin work centers around biochemical factors. But biopolymers catalyzed by clay surfaces, for example, do not necessarily contain any functional (prescriptive) information. How does an algorithmically complex sequence of codons arise in nature which finds phenotypic usefulness only after translation into a completely different language (AA sequence)? How did natural process produce so indirectly the hundreds of needed three-dimensional protein catalysts for life to begin?
Mathematically, it is impossible to go backwards from 20 AA to 64 codons. There is no way to know which of four or six codons, for example, coded a given AA when one tries to go backwards against the "Central Dogma." Prescriptive Information has been lost. Various models of code origin often pursue primordial codon systems of only two nitrogen bases rather than three. At some point, such a two-base codon system must evolve into a three-base codon system. But catastrophic problems such as global frame shifts would have resulted from such a change midstream in the evolution of genetic code.
Environmental selection, if existent at all in a prebiotic environment, is nothing more than after-the-fact differential survivability/reproduction of certain stochastic ensembles in certain environments. How did initial genetic code-certain sequences of codons-come to specify only certain three-dimensional sequences of amino acid strings that "work"?
The winning submission will likely provide both a novel and cardinal conceptual contribution to current biological science and information theory.
The Foundation welcomes theoretical models of a more direct primordial instruction system (one that might have preceded codon transcription and translation) provided the model provides explanation of continuous transition (abiding by the "continuity principle") to current prokaryotic and eukaryotic empirical life.
Inanimate stepping stones of abiotic evolution are essential components to any natural process theory of the molecular evolution of life. Full reign must be given to the exploration of spontaneously forming complexity and to self-ordering inanimate systems. But reductionistic attempts to provide models of life development must not sacrifice the very property of "life" that biology seeks to explain. Coacervates, micelles, vesicles, and various primordial quasimembrane models, for example, may resemble membrane equivalents and merit considerable ongoing research, but should not be confused with the active transport membranes of the simplest known free-living organisms.
Criteria for winning
Major issues
Applicants must provide
A. a well-conceived, detailed hypothetical mechanism explaining how the rise of genetic instructions sufficient to give rise to life as defined in "Definitions" below might have occurred in Nature by natural processes, and an
B. empirical correlation to the real world of biochemistry and molecular biology - not just mathematical or computer models - of how the prescriptive information characteristic of all known living organisms might have arisen.
The mechanism must address four topics:
- The simplest known genome's apparent anticipation and directing of future events toward biological ends, both metabolic and structural;
- The ability of the genome to convey instructions, deliver orders, and actually produce the needed biological end-products;
- The indirectness of recipe-like biological "linguistic" message code - the gap between genotypic prescriptive information (instruction) and phenotypic expression. How did the first genetic instruction arise in its coded format prior to phenotypic realization of progeny from which the environment could select? If a protobiont's genetic code and phenotype were one and the same, how did such a simple system self-organize to meet the nine minimum conditions of "life" enumerated below under "Definitions"? How did stellar energy, the four known forces of physics (strong and weak nuclear forces, electromagnetic force, and gravity), and natural processes produce initial prescriptive information (instruction/recipe) using direct or indirect code?
- The bizarre concentration of singlehanded optical isomers (homochirality of enantiomers) in living things - how did a relatively pure population of left-handed amino acids or right-handed sugars arise out of a chemical environment wherein reactions ordinarily give rise to roughly equal numbers of both right- and left-handed optical isomers?
Definitions
a. By "theory," the Foundation means a thorough explanation and mechanism explaining how natural events might have given rise to a phenomenon like the genetic sign system, algorithmic programming, and code bijection. The Prize is not being offered for creating life in vitro, but for a plausible, empirically supported theory of mechanism within nature.
b. By "mechanism," the Foundation means a scenario of sequential, cause-and-effect (or at least "functionally dependent"), empirically correlated events explaining how genetic prescriptive information (instruction) arose naturally within Nature sufficient to give rise to current life.
c. By "prescriptive information," the Foundation means the instructions necessary for biochemical function, the end-product-oriented specification of monomeric sequence, the "recipe" and "biomessage" of messenger molecules which are so manifest in all known forms of phenomenological life. As pointed out by Hubert Yockey, all known genetic information is observed in a physical matrix sequence that is linear, segregatable, and digital. These linear sequences must be translated via code to other linear sequences for prescriptive information (instruction) to be received at the receiver end of any Shannon channel. They are in effect algorithmic sequences of decision-node configurable switch settings. Algorithms alone produce sophisticated biofunction. A phase space of stochastic ensembles has never been observed to produce even the simplest of biochemical pathways. See "algorithms" under the "Discussion" section.
By "prescriptive information (instruction)," the Foundation does not mean mere order or structure, as in a snowflake. By "prescriptive information (instruction)," the Foundation does not mean mere pattern or periodicity, as in a sine wave, kaleidoscopic image, or redundant inanimate crystal. By "prescriptive information (instruction)," the Foundation does not mean mere physical "complexity," as many complex conglomerates contain no instructional information. By "prescriptive information (instruction)," the Foundation is not merely referring to the probabilistic uncertainty concepts of Shannon, nor to Maxwell-Boltzmann-Gibbs entropy, nor to Kolmogorov-Solomonoff-Chaitin compression theory of mere sequence "complexity" alone. Internal algorithmic compression of alphanumberic symbol sequences defines "complexity." But such comlexity has nothing to do with external algorithmic meaning or function. Sequence complexity is no measure of algorithmic utility. "Function" extends into additional dimensions altogether from mere sequence "complexity." External algorithmic function changes its environment and accomplishes some task external to itself.
d. By "genetic code," the Foundation means "the linguistic-like, symbolic representation of commands from one alphabet and syntactic language (e.g., codon sequence) to another (e.g., AA sequence), conveying seemingly conceptual biological instructions to cell systems." Code is a one-to-one correspondence or "bijection" from one alphabetical system/language to another. See "The source of genetic code in nature" in the "Discussion" section.
Genetic instructions represent a form of "prescriptive information (instruction)" rather than just "Shannon combinatorial/probabilistic information." Genetic "recipe" has been called "aperiodic specified complexity." See "Aperiodic specified complexity" under "Discussion."
e. By sustained, free-living "life," the Foundation means any system which from its own inherent set of biological instructions can perform all nine of the following functions:
1. Delineate itself from its environment through the production and maintenance of a membrane equivalent, most probably a rudimentary or quasi-active-transport membrane necessary for selective absorption of nutrients, excretion of wastes, and overcoming osmotic and toxic gradients,
2. Write, store, and pass along into progeny prescriptive information (instruction) needed for organization; provide instructions for energy derivation and for needed metabolite production and function; symbolically encode and communicate functional message through a transmission channel to a receiver/decoder/destination/effector mechanism; integrate past, present and future time into its biological prescriptive information (instruction) content,
3. Bring to pass the above recipe instructions into the production or acquisition of actual catalysts, coenzymes, cofactors, etc.; physically orchestrate the biochemical processes/pathways of metabolic reality; manufacture and maintain physical cellular architecture; establish and operate a semiotic system using "signal molecules"
4. Capture, transduce, store, and call up energy for utilization (work),
5. Actively self-replicate and eventually reproduce, not just passively polymerize or crystallize; pass along the apparatus and "know-how" for homeostatic metabolism and reproduction into progeny,
6. Self-monitor and repair its constantly deteriorating physical matrix of bioinstruction retention/transmission, and of architecture,
7. Develop and grow from immaturity to reproductive maturity,
8. Productively react to environmental stimuli. Respond in an efficacious manner that is supportive of survival, development, growth, and reproduction, and
9. Possess relative genetic stability, yet sufficient diversity to allow for adaptation and potential evolution.
All classes of archaea, bacteria, and every other known free-living organism, meet all nine of the above criteria. Eliminate any one of the above nine requirements, and it remains to be demonstrated whether that system could remain "alive."
RNA strands, DNA strands, prions, viroids, and viruses shall not be considered free-living organisms, since they fail to meet many of the above well-recognized characteristics of independent "life."
Even in historical science, there must be some degree of empirical accountability to our theories. Proposing a mechanism that explains the origin of life must not consist of "defining down" the meaning and essence of the observable phenomenon of "life" to include "nonlife" in order to make our theories "work." Any scientific life-origins theory must connect with "life" as we observe it (the "continuity principle").
Science will never be able to abandon its empirical roots in favor of purely theoretical conjecture. On the other hand, science must constantly guard itself against Kuhnian paradigm ruts. We must be open-minded to the possibility that life has not always taken the form that we currently observe. We must take into consideration the problems inherent in any historical science where the observation of past realities is impossible.
Biophysicist Hubert P. Yockey makes the unique observation that "there is nothing in the physico-chemical world [apart from life] that remotely resembles reactions being determined by a sequence and codes between sequences. The existence of a genome and the genetic code divides living organisms from non-living matter." (Computers and Chemistry, 24 (2000) 105-123). This may well constitute the most concise and parsimonious dichotomization of animacy from inanimacy available in the literature.
We must remember, however, that the full compliment of nucleic acid code, ribozymes, and protein enzymes are still present immediately after cell death. Life, therefore, would appear not to be reducible to coded prescriptive information (instruction) alone. Life is also not "a bag of enzymes." "Life" is characterized by ongoing homeostatic metabolic process and algorithmic function, including development, growth, and reproductive potential. The inability of mules to reproduce has no relevance to discussions of protocellular viability.
Discussion
1. Entropy
By "entropy" as it relates to information theory, the Foundation adopts Hubert P. Yockey's distinction between Maxwell-Boltzmann-Gibbs entropy, Shannon probability-distribution entropy, and Kolmogorov-Solomonoff-Chaitin sequence/algorithmic complexity. (See Information Theory and Molecular Biology, Cambridge University Press, 1992, sections 2.2.2 and 2.4.1 - 2.4.6). (See also, Yockey, H.P., (1974) "An application of information theory to the Central Dogma and the sequence hypothesis." Journal of Theoretical Biology, 46, 369-406, and Yockey, H.P.(1981) Self Organization, Origin of Life Scenarios, and Information Theory, J. Theor. Biology, 91, 13-31, and Yockey, H.P. (2000) Origin of life on earth and Shannon's theory of communication, Comput Chem, 24, 1, pp 105-123) Yockey argues that there is no "balancing act" between algorithmic informational entropy and Maxwell-Boltzmann-Gibbs-type entropy. The two are not on the same see-saw. The two probability spaces are not isomorphic. Information theory lacks the integral of motion present in thermodynamics and statistical mechanics. In addition, there is no code linking the two "alphabets" of stochastic ensembles. Kolmogorov-Solomonoff-Chaitin complexity does not reside in the domain of stochastic ensembles of statistical mechanics. They have no relation despite endless confusion and attempts in the literature to merge the two.
"Highly ordered" is paradoxically opposite from "complex" in algorithmic-based information theory. The emergent property of "instructions," "organization," and the "message" of "messenger biomolecules" is simply not addressed in Maxwell-Boltzmann-Gibbs equations of heat equilibration and energy flux between compartments. Surprisingly, the essence of genetic "prescriptive information" and "instructions" is not addressed by current "information theory" either. Shannon information theory concerns itself primarily with data transmission, reception, and noise-reduction processing without regard for the essence of the "message" itself.
The Foundation questions whether "order," physical "complexity," or "shared entropy" are synonymous with "prescriptive information," "instructions," or "organization."
Christoph Adami emphasizes that information is always "about something, and cannot be defined without reference to what it is information about." It is "correlation entropy" that is "shared" or "mutual." Thus, says Adami, "Entropy can never be a measure of complexity. Measuring correlations within a sequence, like Kolmogorov and Chaitin (and Lempel-Ziv, and many others) is not going to reveal how that sequence is correlated to the environment within which it is to be interpreted. Information is entropy "shared with the world," and the amount of information a sequence shares with its world represents its complexity." (Personal communication; see also PNAS, April 25, 2000, 97, #9, 4463-4468).
Differences of perspective among information theorists are often definitional. "Complexity" and "shared entropy" (shared uncertainty between sender and receiver) has unfortunately often been used synonymously with "prescriptive information (instruction)." But is it? Mere complexity and shared entropy seem to lack the specification and orchestrational functionality inherent in the genetic "instruction" system of translation.
The confusion between algorithmic instruction and Maxwell-Boltzmann-Gibbs entropy may have been introduced through the thought experiment imagining Maxwell's Demon - a being exercising intelligent choice over the opening and closing of a trap door between compartments. Statistical mechanics has no empirical justification for the introduction of purposeful control over the trap door.
Solar energy itself has never been observed to produce prescriptive information (instruction/organization). Photons are used by existing instructional mechanisms which capture, transduce, store, and utilize energy for work. Fiber optics is used by human intelligence to transmit meaningful prescriptive information (instruction) and message. But raw energy itself must not be confused with functional prescriptive information/instructions. The latter is a form of algorithmic programming. Successions of certain decision-node switch settings determine whether a genetic "program" will "work" to accomplish its task.
2. Is life autonomous?
Some argue that life exhibits the characteristics of autonomy. Life is orchestrated by the prescriptive information (instruction) content and the inherent systems with which it finds itself. It is directed (passive voice) by its inherited genome. Because of this others argue that even a prokaryote's autonomy is apparent rather than real. Cells are fully dependent upon "recipe" and control mechanisms delivered from a prior and external source. Cells also remain dependent upon their environment, especially for energy. But without the transducing mechanisms instructed by its genome, energy will only accelerate its demise.
3. Does life display Negentropy?
1. It is mathematically impossible for entropy to be a negative entity. (Yockey, 1992, Information Theory and Molecular Biology, Cambridge University Press, p 84)
2. Organisms do not violate the Second Law of Thermodynamics any more than any other physical entity in open systems far from equilibrium. Their existing genetic instructions, command and control mechanisms, and machinery simply allow them to process incoming nutrients and energy in full accord with the Second Law. The problem lies in the derivation of the functional biological information that makes both of these processes possible. By what mechanism did initial instructions arise sufficient to produce such highly conceptual metabolic and replicative systems? This is the quest of The Gene Emergence Project, and the object of The Origin-of-Life Prize offer.
4. Specified aperiodic complexity
Many investigators, such as Leslie Orgel (Origins of Life, 1973, New York, John Wiley, 189-190), for many years have regarded genetic information as having an additional component besides mere complexity. Matrices of prescriptive information retention are not only extremely improbable ensembles, but ones which are specified in a way that yields biofunction. "Specified complexity" means that only a certain few out of a large sample space of potential or real ensembles will produce metabolic function. Specified complexity instructs and integrates biochemical pathways into homeostatic metabolism.
5. Algorithmic instruction
Life is an integration of many algorithmic processes that give rise to biofunction and overall homeostatic metabolism. Algorithms are processes or procedures that give rise to useful function. Algorithms are not merely linear sequences of symbols. They do something. Each symbol represents a choice from among symbol options. Each choice is critical to the determination of eventual function. There is an organizational property - a certain element of seeming conceptuality - to the biological information /instructions that produce the citric acid cycle, for example. This aspect raises bioinformation to a more instructive, orchestrational, and recipe level than mere physical order, complexity, probabilistic uncertainty, or "mutual uncertainty" between two sets.
Algorithmic compression schemes are valuable in defining plain complexity. Such algorithms address internal sequence compressibility only. But these types of algorithms tell us nothing about whether the sequence instructs any function external to itself. The degree of compressibility is not critical to defining prescriptive information (instruction). The functionality the sequence produces is.
Algorithms are usually schema of successive decision node "choices" that lead somewhere useful. The sequence of choices accomplishes some useful task. Each decision node represents a fork in the road. One false turn, and the potential function at the end of the sequence can be lost. Dendrograms of these decision node choice options give rise to many orders of magnitude of potential terminal tree branches. Only a few of these branches usually yield sophisticated function. A biopolymer represents this sequence of decision-node symbol "choices." Even when homologous protein sequences are considered from genetic drift, sometimes the equivalent of only one branch out of 2^30 (10^10) branches in sequence space "work" as the needed enzyme. Life-origin scenarios must provide explanation and mechanism for how such unlikely algorithmic strings come together at the same place and at the same time to produce not only the local function of each individual algorithmic program, but the integration of many hundreds of such strings into homeostatic metabolism.
6. The source of genetic information in nature
No theory of genetic information is complete without a model of mechanism for the source of such prescriptive information within Nature. It is not sufficient for a submission to the Prize to limit discussion of prescriptive information (instruction) theory to its replication, transmission, modification, or matrix of information retention. All submissions must address the source of the prescriptive information through non-supernaturalistic natural processes. Which of the four known forces of physics, or what combination of these forces, produced prescriptive, functional information, and how? What is the empirical evidence for this kind of prescriptive information (instruction) spontaneously arising within Nature?
7. Genetic code
In all known phenomenological life, genetic code manifests the conveyance of a functional coded message, using a sign system, to distant sites through an information channel to energy-consuming decoding receivers - ribosomal "machines,"
• symbolic, indirect representation of that message from one alphabet into another (e.g., codons of nitrogen base "language" being translated into the end-product of physical amino acid sequence "language.")
• prespecification of extremely unlikely and complex future events (see Dembski in suggested readings below) suggesting "apparent intent," "apparent planning," or "apparent purpose." (as Richard Dawkins describes it, "apparent design"),
• instructions capable of effecting and affecting many individual manufacturing processes, and of mediating the cooperation of all of those diverse processes toward the one organismal and seemingly "conceptual" end of being and staying alive, and
• the ability of that information (instruction) not only to give the directions or orders of what should be done, but to bring to pass those orders in the form of actual physical molecules, products, and life processes.
• the seemingly "irreducible complexity" argued by Michael Behe (see suggested readings below)
• the initial writing of this prescriptive information by nature, not just the modification of existing genetic instruction through mutation.
8. Scaffolding models
a. "Scaffolding" models of prelife (e.g., silicone/clay matrix models) constitute acceptable submissions as long as detailed and plausible hypothetical mechanisms along with empirical correlation are provided linking such inanimate crystal matrix models to "the arch" of current carbon-chemistry life. Such models would have to explain how random defects in crystal layers got arranged into functional genetic prescriptive information (instruction). The issue is not the medium or matrix that retains the information. The issue is the source of prescriptive information (instruction) itself in any medium or matrix of Nature.
b. "Scaffolding" models would also have to explain how inanimate crystals or the ions/radicals adsorbed onto them progressively acquired the nine minimum functions and capabilities of living organisms listed under the provided definition of "life."
9. Biochemical correlation
a. The hypothetical mechanism must demonstrate correspondence with "the real world" of biochemistry.
b. The submission must provide adequate empirical support strongly suggesting that such a hypothetical scenario can take place naturally in a prebiotic environment. Simulation of abiogenesis must be independent of the factor of human intelligence that is so often subconsciously incorporated into computer hardware/software experimental design and simulation.
c. Thermodynamic realities must be clearly addressed, including specific discussion of any supposed pockets of momentary exception to the Second Law of increasing Maxwell-Boltzmann-Gibbs entropy. The Foundation's view is that Prigogine's dissipative structures, and life itself, operate within the constraints of the 2nd Law.
Maxwell-Boltzmann-Gibbs entropy must not be confused with statistical Shannon entropy or Kolmogorov-Chaitin-Solomonoff-Yockey "complexity." The latter two are nonphysical, abstract, mathematical constructs. All physical matrices of prescriptive information retention, however, are subject to the plight of Maxwell-Boltzmann-Gibbs entropy. They manifest a tendency toward deterioration in both closed and open systems. Repair mechanisms for these messenger biomolecules, therefore, require all the more starting instructional integrity. Prescriptive information would have been necessary in any primordial life form's genome to correct for continuous noise corruption of its functional protogenes. Deterioration of existing recipe in a physical matrix is far more probable than the spontaneous writing of new conceptually complex metabolic algorithms. Building-block synthesis, for instance, would have required something like a reductive citric acid cycle. There are no simple algorithms for integrating such a multistep, highly-directional pathway.
d. Empirical support does not have to be original research, but can be gleaned from existing scientific literature. Previously published empirical support must be described in detail and well referenced within the applicant's published research paper, explaining exactly how those controlled observations demonstrate empirical correlation with the applicant's theory.
10. "Design" anthropomorphisms
It is easy for us to attribute "apparent instruction" or "apparent design" to projections of human intelligence onto the data. Based on current knowledge of molecular biology, any protocell or protobiont imaginable clearly must have manifested such "concept" to come to life long before any humans appeared on the scene to project or anthropomorphize anything. The Foundation believes that use of the words "order" and "complexity" are grossly inadequate euphemisms for the clearly observable prescriptive information, genetic instructions, biomessage, and functional biochemical pathways inherent in the simplest free-living organisms. The Foundation further believes that these empirical properties of genetic instruction and life can be investigated scientifically.
The simplest known living organisms are replete with empirical evidence of organizational unity and coherence which directs future biochemical events toward undeniable ends and purposes. Prokaryotes such as Archaea exhibit the integration of multiple biological systems into extraordinary organismic cooperation. The Foundation interprets such observable genetic instructions to be undeniable empirical evidence of objectively existent "concept" independent of human mentation. The "unusual effectiveness of mathematics" in physics offers more evidence. Such conceptual capacity predated human intelligence in any evolutionary paradigm. A hypothetical mechanism is therefore needed for two aspects of life origin:
• how seemingly unintelligent natural processes could have written such highly prescriptive recipe/message linguistic-like code, and
• how such an indirect system could have arisen which effects (brings into existence) so many hundreds of integrated and far-removed phenotypic processes and products in the simplest organisms.
11. Appeals to unknown laws
Appealing to unknown "laws" as the source of biological instruction constitutes a "category error" of logic theory. "Laws" do not cause anything. They are merely human generalizations, mental constructions, and mathematical descriptions of existing forces and mass-energy relationships. Even "chance" is a probabilistic rational construct. Neither chance nor "laws" cause effects. Unknown laws, therefore, cannot provide a mechanism for prescriptive information (instruction) genesis. Appealing to unknown laws constitutes a "naturalism of the gaps," corresponding to supernaturalists' appealing to a "God of the gaps" for scientific explanation. Neither is acceptable in naturalistic science.
12. Infinity issues
Appeals to multiple or "parallel" cosmoses or to an infinite number of cosmic "Big Bang/Crunch" oscillations as essential elements of proposed mechanisms are not acceptable in submissions due to a lack of empirical correlation and testability. Such beliefs are without hard physical evidence and must therefore be considered unfalsifiable, currently outside the methodology of scientific investigation to confirm or disprove, and therefore more mathematically theoretical and metaphysical than scientific in nature. Recent cosmological evidence also suggests insufficient mass for gravity to reverse continuing cosmic expansion. The best cosmological evidence thus far suggests the cosmos is finite rather than infinite in age.
13. Computerization
a. Computerized models must be free of subtle, inherent teleological design flaws which become incorporated into the model itself. The insidious role of human intelligence in both hardware and software must be acknowledged, addressed, and somehow divorced from hypothetical models themselves.
b. Models based on conditional probabilities must justify empirically why the environment would have selected for each plateau along the way. For example, why would the environment have favored and preserved the intermediary steps in many metabolic pathways of archaea when no useful product is produced until the last step? Many of these multistep, indirect manufacturing pathways constitute all-or-none processes. Such biochemical pathways have no phenotypic "plateaus" in physical biochemical reality to support theoretical arguments of selectable incremental function. Yet the random occurrence of the full pathway as a whole is statistically prohibitive in a trillion billion years, let alone in the mere 15-billion-year age of our cosmos.
c. Other factors limit the number of statistical trials for an exploding cosmic egg to randomly give rise to such sophisticated pathways in 15-billion years. The finite number of nucleons available in the cosmos to react with one another (10^80?) is an example. We can no longer appeal to infinity of particles, space, or time as a means of overcoming the statistical prohibitiveness within the only cosmos with which we have empirical experience. Abundant data, mathematical proofs, and our best theories all suggest that our cosmos is finite, not infinite. We have no scientific knowledge of any other cosmos, let alone an infinite number of imaginary cosmoses.
d. Parallel computer models must similarly have direct empirical correlation with naturally occurring environmental, chemical, biochemical, and molecular biological scenarios. "Directed evolution" experiments must not incorporate the artificial selection of investigators into their experimental design. "Directed evolution" is a self-contradictory phrase. Evolution by definition is never directed. Evolution in fact has no goal or purpose.
Bibliografia recomendada:
Schroedinger, E. What is Life? (Cambridge, Cambridge University Press, 1955)
Donald M. MacKay. Information, Mechanism and Meaning (Cambridge, MA: M.I.T. Press, 1969)
Jacques Monod. Chance and Necessity (New York: Knopf, 1971)
Leslie E. Orgel. The Origins of Life: Molecules and Natural Selection (New York, John Wiley, 1973)
Stanley L. Miller and Leslie E. Orgel. The Origins of Life on Earth (Eaglewood Cliffs: Prentice-Hall, 1974)
R. W. Hamming. Coding and Information Theory (Englewood Cliffs, Prentice- Hall, l980)
M. Kimura. The Neutral Theory of Molecular Evolution (Cambridge: Cambridge University Press, 1983)
Charles B. Thaxton, Walter L. Bradley, Roger L. Olsen. The Mystery of Life's Origin (Dallas: Lewis and Stanley, 1984)
A. G. Cairns-Smith. Seven Clues to the Origin of Life (New York: Cambridge University Press, 1985).
Freeman Dyson. The Origins of Life (Cambridge: Cambridge University Press, 1985) Now second edition, (1999).
Robert Shapiro. Origins: A Skeptic's Guide to the Creation of Life on Earth (New York: Summit Books, 1986)
John Maynard-Smith. The Problems of Biology (Oxford: Oxford University Press, 1986)
Bernd-Olaf Küppers. Information and the Origin of Life (Cambridge: MIT Press, 1990)
Cyril Ponnamperuma and F. R. Eirich, Editors Prebiological Self-Organization of Matter (A. Deepak Publishing: Hampton, VA, 1990)
Christian De Duve, Blueprint for a Cell:The Nature and Origin of Life (Burlington, NC: Patterson, 1991)
Hubert P. Yockey. Information Theory and Molecular Biology, (Cambridge: Cambridge University Press, 1992).
H. J. Morowitz. Beginnings of Cellular Life: Metabolism Recapitulates Biogenesis (New Haven, CT: Yale University Press, 1992)
Cyril Ponnamperuma and Julian Chela-Flores. Chemical Evolution: Origin of Life Proceedings of The Trieste Conference on Chemical Evolution and the Origin of Life, 26-30 October, 1992 (Hampton, VA: A Deepak Publishing, 1993)
Walter James ReMine. The Biotic Message (Saint Paul, MN: St. Paul Science, 1993)
Stuart A. Kauffman. The Origins of Order: Self-Organization and Selection in Evolution (New York: Oxford University Press, 1993)
David W. Deamer and Gail R. Fleischaker. Origins of Life: The Central Concepts (Boston: Jones and Bartlett Publishers, 1994).
Periannan Senapathy. Independent Birth of Organisms (Madison: Genome Press, 1994).
Julian Chela-Flores, Mohindra Chadha, Alicia Negron-Mendoza, Tairo Oshima, Editors. Chemical Evolution: Self-organization of the Macromolecules of Life Proceedings of The Trieste Conference on Chemical Evolution and the Origin of Life, 25-29 October 1993 (Hampton, VA: A. Deepak Publishing, 1995)
John H. Holland. Hidden Order: How Adaptation Builds Complexity (Reading, Mass.: Perseus Books, 1995)
Christopher G. Langton. Artificial Life: An Overview (Cambridge, MA: MIT Press, 1995)
Stuart Kauffman. At Home in the Universe: The Search for the Laws of Self-Organization and Complexity (New York: Oxford University Press, 1995).
Christian De Duve, Vital Dust--Life as a Cosmic Imperative (New York: Basic Books, 1995)
Syozo Osawa, Evolution of the Genetic Code (Oxford: Oxford University Press, 1995)
Lynn Margulis and D. Sagan. 'What is Life' (London: Weidenfeld and Nicholson, 1995)
Rizzoti (ed.): 'Defining Life' (Padua, Italy: University Padua Press, 1996)
Geoffrey Zubay. Origins of Life on the Earth and in the Cosmos (New York: WCB/McGraw Hill, 1996)
Michael J. Behe. Darwin's Black Box, (New York: The Free Press/Simon and Schuster, 1996).
Julian Chela-Flores and Francois Raulin. Chemical Evolution: Physics of the Origins and Evolution of Life, Proceedings of the 4th Trieste Conference on Chemical Evolution, Trieste, Italy, 4-8 September 1995 (Netherlands: Kluwer Academic Publishers, 1996)
Manfred Eigen and Ruthild Winkler-Oswatitsch. Steps Toward Life: A Perspective on Evolution (Oxford: Oxford University Press, 1996)
Dean Overman. The Case Against Accident and Self-Organization, (New York: Rowman and Littlefield, 1997)
Christoph Adami. Artificial Life (New York: Springer-Telos, 1998)
William A. Dembski. The Design Inference, in Cambridge Studies in Probability, Induction, and Decision Theory. (New York: Cambridge University Press, 1998).
Noam Lahav. Biogenesis: Theories of Life's Origin (Oxford: Oxford University Press, 1998)
André Brack. The Molecular Origins of Life: Assembling the Pieces of the Puzzle (New York: Cambridge University Press, 1998)
Michael Gross. Life on the Edge (New York: Plenum Press, 1998)
John Maynard Smith. Shaping Life: Genes, Embryos, and Evolution (UK, Weidenfeld & Nicholson, 1998; U.S., Yale University Press, 1999)
James P. Ferris, Editor. Origins of Life and Evolution of the Biosphere, Papers presented at the 1996 ISSOL Meeting in Orleans, France. Volume 28, Nos.4-6 October 1998 (Boston: Kluwer Academic Publishers, 1998)
Martin J. Medhurst and John Angus Campbell, Editors Rhetoric & Public Affairs (East Lansing, Michigan: Michigan State University Press, Vol 1, No 4, Winter 1998 Entire issue)
Fred Hoyle. Mathematics of Evolution (Memphis, Tenn: Acorn Enterprises, 1999)
John Maynard Smith and Eors Szathmary. The Origins of Life: From the Birth of Life to the Origins of Language (Oxford: Oxford University Press, 1999)
Gesteland, R. F., Cech, T. R. & J F. Atkins, eds. The RNA World (Plainview, 2ND Edition, (NY: Cold Spring Harbor Lab. Press, 1999)
Werner R. Loewenstein. The Touchstone of Life: Molecular Information, Cell Communication, and the Foundations of Life (Oxford: Oxford University Press, 1999)
Hans Kuhn and Horst-Dieter Forsterling. Principles of Physical Chemistry (England: Wiley Press, 1999) pp 880-921; see also p 953
Gyula Palyi, Claudia Zucchi, Luciano Caglioti (Eds.) Advances in BioChirality., (Amsterdam: Elsevier, 1999)
Freeman Dyson. The Origins of Life (Cambridge: Cambridge University Press, Second Edition, (1999)
David Berlinski. The Advent of the Algorithm: the Idea That Rules the World (New York: Harcourt, Inc., 2000)
Iris Fry. The Emergence of Life on Earth: A Historical and Scientific Overview (N.J.: Rutgers University Press, 2000)
L Kay. Who Wrote the Book of Life? A History of the Genetic Code (Stanford: Stanford Univeristy Press, 2000)
Richard Sole and Brian Goodwin. Signs of Life: How Complexity Pervades Biology (New York: Basic Books Persius, 2000)
Stuart Kauffman. Investigations (New York: Oxford University Press, 2000)
Christopher Wills and Jeffrey Bada. The Spark of Life, (Cambridge, Massachusetts: Persius, 2001)
Gyula Palyi, Claudia Zucchi, Luciano Caglioti (Eds.) Fundamentals of Life, (Paris: Elsevier, 2002)
William A. Dembski, No Free Lunch, (New York, Rowman and Littlefield, 2002)
Henry Harris, Things Come to Life: Spontaneous generation Revisited, (Oxford, Oxford University Press, 2002)
J William Schopt, Ed, Life's Origin: The Beginnings of Biological Evolution, (Ewing, N. J., Univer. of California Press, 2002)
Tom Fenchel, Origin and Early Evolution of Life (Oxford, Oxford University Press, 2003)
Marcello Barbieri, The Organic Codes: An Introduction to Semantic Biology (Cambridge, UK, Cambridge University Press, 2003)
Tibor Ganti, The Principles of Life (Oxford, UK, Oxford Unversity Press, 2003)
Nancy Forbes, Imitation of Life: How Biology is Inspiring Computing (MIT Press, 2004)
Clive Trotman, The Feathered Onion - Creation of life in the Universe (John Wiley and Sons, 2004)
Hubert P. Yockey, Information Theory, Evolution, and the Origin of Life (Cambridge, Cambridge University Press, 2004)
Robert M. Hazen, Genesis: The Scientific Quest for Life's Origin (Joseph Henry Press, Washington, D.C.2005)
Marc W. Kirschner and John C. Gerhart: The Plausibility of Life: Resolving Darwin's Dilemma (Yale University Press, New Haven, 2005)
 Nas pesquisas que realizo atualmente sobre a controvérsia entre Mivart e Darwin, tenho achado algumas coisas muito interessantes sobre Darwin e os demais atores. Há, contudo, uma declaração de C.D. Darlington, geneticista evolucionista, que me deixou e deixa bastante intrigado: "... há um sentimento natural (entre os cientistas) de que uma das maiores de nossas figuras [Darwin] não deva ser dissecada, pelo menos por um de nós". [1] Por que Darwin tem que ser dissecado por outros fora da Biologia? Qual a razão de 'blindar' Darwin de quaisquer críticas?
Nas pesquisas que realizo atualmente sobre a controvérsia entre Mivart e Darwin, tenho achado algumas coisas muito interessantes sobre Darwin e os demais atores. Há, contudo, uma declaração de C.D. Darlington, geneticista evolucionista, que me deixou e deixa bastante intrigado: "... há um sentimento natural (entre os cientistas) de que uma das maiores de nossas figuras [Darwin] não deva ser dissecada, pelo menos por um de nós". [1] Por que Darwin tem que ser dissecado por outros fora da Biologia? Qual a razão de 'blindar' Darwin de quaisquer críticas? Depois do que li no blog Uncommon Descent o artigo "Was Blyth the true scientist and Darwin merely a plagiarist and charlatan?", fiquei com uma baita pulga na orelha – seria Darwin um reles charlatão plagiador? Seria Edward Blyth quem pensou primeiro sobre a "seleção natural"? Darwin 'roubou' a idéia de Blyth, um criacionista?
Depois do que li no blog Uncommon Descent o artigo "Was Blyth the true scientist and Darwin merely a plagiarist and charlatan?", fiquei com uma baita pulga na orelha – seria Darwin um reles charlatão plagiador? Seria Edward Blyth quem pensou primeiro sobre a "seleção natural"? Darwin 'roubou' a idéia de Blyth, um criacionista?