
A estabilidade em larga escala dos cromossomos
Large Scale Chromosome Folding Is Stable against Local Changes in Chromatin Structure
Ana-Maria Florescu , Pierre Therizols, Angelo Rosa
Published: June 13, 2016 http://dx.doi.org/10.1371/journal.pcbi.1004987

Abstract
Characterizing the link between small-scale chromatin structure and large-scale chromosome folding during interphase is a prerequisite for understanding transcription. Yet, this link remains poorly investigated. Here, we introduce a simple biophysical model where interphase chromosomes are described in terms of the folding of chromatin sequences composed of alternating blocks of fibers with different thicknesses and flexibilities, and we use it to study the influence of sequence disorder on chromosome behaviors in space and time. By employing extensive computer simulations, we thus demonstrate that chromosomes undergo noticeable conformational changes only on length-scales smaller than 105 basepairs and time-scales shorter than a few seconds, and we suggest there might exist effective upper bounds to the detection of chromosome reorganization in eukaryotes. We prove the relevance of our framework by modeling recent experimental FISH data on murine chromosomes.
Author Summary
A key determining factor in many important cellular processes as DNA transcription, for instance, the specific composition of the chromatin fiber sequence has a major influence on chromosome folding during interphase. Yet, how this is achieved in detail remains largely elusive. In this work, we explore this link by means of a novel quantitative computational polymer model for interphase chromosomes where the associated chromatin filaments are composed of mixtures of fibers with heterogeneous physical properties. Our work suggests a scenario where chromosomes undergo only limited reorganization, namely on length-scales below 105 basepairs and time-scales shorter than a few seconds. Our conclusions are supported by recent FISH data on murine chromosomes.
Citation: Florescu A-M, Therizols P, Rosa A (2016) Large Scale Chromosome Folding Is Stable against Local Changes in Chromatin Structure. PLoS Comput Biol 12(6): e1004987. doi:10.1371/journal.pcbi.1004987
Editor: Marc A. Marti-Renom, CNAG - Centre Nacional d’Anàlisi Genòmica and CRG - Centre de Regulació Genòmica, SPAIN
Received: November 5, 2015; Accepted: May 17, 2016; Published: June 13, 2016
Copyright: © 2016 Florescu et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original author and source are credited.
Data Availability: All relevant data are within the paper and its Supporting Information files.
Funding: AR has received financing from Italian Ministry of Research through project PRIN - 2010 HXAW77 (http://prin.miur.it/). PT has recieved financing from Agence Nationale de la Recherche through grant IDEX-SLI (DXCAIHUSLI-EF14). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Competing interests: The authors have declared that no competing interests exist.
FREE PDF GRATIS: PLoS Computational Biology
Singularidade de coordenação de corrida humana: a integração de inovações evolutivas modernas e antigas
MINI REVIEW ARTICLE
Front. Psychol., 11 April 2016 | http://dx.doi.org/10.3389/fpsyg.2016.00262
Uniqueness of Human Running Coordination: The Integration of Modern and Ancient Evolutionary Innovations
John Kiely1,2* and David J. Collins1
1School of Health and Wellbeing, Institute of Coaching and Performance, University of Central Lancashire, Preston, UK
2Fitness Department, Irish Rugby Football Union, Dublin, Ireland

Abstract
Running is a pervasive activity across human cultures and a cornerstone of contemporary health, fitness, and sporting activities. Yet for the overwhelming predominance of human existence running was an essential prerequisite for survival. A means to hunt, and a means to escape when hunted. In a very real sense humans have evolved to run. Yet curiously, perhaps due to running's cultural ubiquity and the natural ease with which we learn to run, we rarely consider the uniqueness of human bipedal running within the animal kingdom. Our unique upright, single stance, bouncing running gait imposes a unique set of coordinative difficulties. Challenges demanding we precariously balance our fragile brains in the very position where they are most vulnerable to falling injury while simultaneously retaining stability, steering direction of travel, and powering the upcoming stride: all within the abbreviated time-frames afforded by short, violent ground contacts separated by long flight times. These running coordination challenges are solved through the tightly-integrated blending of primitive evolutionary legacies, conserved from reptilian and vertebrate lineages, and comparatively modern, more exclusively human, innovations. The integrated unification of these top-down and bottom-up control processes bestows humans with an agile control system, enabling us to readily modulate speeds, change direction, negotiate varied terrains and to instantaneously adapt to changing surface conditions. The seamless integration of these evolutionary processes is facilitated by pervasive, neural and biological, activity-dependent adaptive plasticity. Over time, and with progressive exposure, this adaptive plasticity shapes neural and biological structures to best cope with regularly imposed movement challenges. This pervasive plasticity enables the gradual construction of a robust system of distributed coordinated control, comprised of processes that are so deeply collectively entwined that describing their functionality in isolation obscures their true irrevocably entangled nature. Although other species rely on a similar set of coordinated processes to run, the bouncing bipedal nature of human running presents a specific set of coordination challenges, solved using a customized blend of evolved solutions. A deeper appreciation of the foundations of the running coordination phenomenon promotes conceptual clarity, potentially informing future advances in running training and running-injury rehabilitation interventions.
FREE PDF GRATIS: Frontiers in Psychology
Evolução do código genético através da quebra de simetria progressiva
Volume 347, 21 April 2014, Pages 95–108
Evolution of the genetic code through progressive symmetry breaking
- Route Cantonale 103, Saint Sulpice VD, Switzerland
- Received 12 December 2012, Revised 18 December 2013, Accepted 1 January 2014, Available online 14 January 2014
- doi:10.1016/j.jtbi.2014.01.002
- Abstract
- Evolution of the genetic code in an early RNA world is dependent on the steadily improving specificity of the coevolving protein synthesis machinery for codons, anticodons, tRNAs and amino acids. In the beginning, there is RNA but the machinery does not distinguish yet between the codons, which therefore all encode the same information. Synonymous codons are equivalent under a symmetry group that exchanges (permutes) the codons without affecting the code. The initial group changes any codon into any other by permuting the order of the bases in the triplet as well as by replacing the four RNA bases with each other at every codon position. This group preserves the differences between codons, known as Hamming distances, with a 1-distance corresponding to a single point mutation. Stepwise breaking of the group into subgroups divides the 64 codons into progressively smaller subsets – blocks of equivalent codons under the smaller symmetry groups, with each block able to encode a different message. This formalism prescribes how the evolving machinery increasingly differentiates between codons. The model indicates that primitive ribosomes first identified a unique mRNA reading frame to break the group permuting the order of the bases and subsequently enforced increasingly stringent codon–anticodon basepairing rules to break the subgroups permuting the four bases at each codon position. The modern basepairing rules evolve in five steps and at each step the number of codon blocks doubles. The fourth step generates 16 codon blocks corresponding with the 16 family boxes of the standard code and the last step splits these boxes into 32 blocks of commonly two, but rarely one or three, synonymous codons. The evolving codes transmit at most one message per codon block and as the number of messages increases so does the specificity of the code and of protein synthesis. The selective advantage conferred by better functioning proteins drives the symmetry breaking process. Over time paralogous tRNA evolution expands the anticodon repertoire, which is divided into anticodon blocks matching the codon blocks under the stage-specific ribosomal basepairing rules. Contemporaneously an expanding family of primitive aminoacyl-tRNA synthetases (aaRSs) divides the tRNA diversities into various different and overlapping subsets: each aaRS accepts some tRNAs but rejects all others and several aaRSs may accept the same tRNA species. Selection favoring less ambiguous codes eliminates these overlaps and also imposes the ribosomal anticodon block division as ambiguity arises when different aaRSs accept tRNAs of the same anticodon block. Only when the tRNAs of one or several anticodon blocks are accepted by a unique aaRS does the code become specific. This coding pattern is observed in the standard code and the evolution of amino acid assignments by primitive aaRSs onto tRNAs is traced back via tRNA trees that picture a gradual division of tRNA diversities into blocks with increasingly specific amino acid assignments. Symmetry breaking combined with continuous selection for codes carrying more information evolves increasingly specific codes and efficiently traverses an immense space of all possible codes (>1084) to give rise to the standard code.
Keywords
- Aminoacyl-tRNA synthetase;
- Codon graph;
- Hamming distance;
- Ribosome;
- Shannon entropy
- FREE PDF GRATIS: Journal of Theoretical Biology

Eutérios experimentaram elevadas taxas evolutivas no período imediato da extinção em massa do Cretáceo-Paleogeno
quarta-feira, junho 29, 2016
Eutherians experienced elevated evolutionary rates in the immediate aftermath of the Cretaceous–Palaeogene mass extinction
Thomas John Dixon Halliday, Paul Upchurch, Anjali Goswami
Published 29 June 2016. DOI: 10.1098/rspb.2015.3026
Abstract
The effect of the Cretaceous–Palaeogene (K–Pg) mass extinction on the evolution of many groups, including placental mammals, has been hotly debated. The fossil record suggests a sudden adaptive radiation of placentals immediately after the event, but several recent quantitative analyses have reconstructed no significant increase in either clade origination rates or rates of character evolution in the Palaeocene. Here we use stochastic methods to date a recent phylogenetic analysis of Cretaceous and Palaeocene mammals and show that Placentalia likely originated in the Late Cretaceous, but that most intraordinal diversification occurred during the earliest Palaeocene. This analysis reconstructs fewer than 10 placental mammal lineages crossing the K–Pg boundary. Moreover, we show that rates of morphological evolution in the 5 Myr interval immediately after the K–Pg mass extinction are three times higher than background rates during the Cretaceous. These results suggest that the K–Pg mass extinction had a marked impact on placental mammal diversification, supporting the view that an evolutionary radiation occurred as placental lineages invaded new ecological niches during the Early Palaeocene.
FREE PDF GRATIS: Proc R Soc B
Taxas de mutação e a evolução da estrutura da linha germinativa
Mutation rates and the evolution of germline structure
Aylwyn Scally
Published 20 June 2016. DOI: 10.1098/rstb.2015.0137

Cellular genealogies in a two-offspring family. Solid black lines represent cellular lineages; germ cell populations are shaded green (in females) or blue (in males), somatic cells are shaded orange. Darker somatic populations represent the cellular ancestors of somatic cells sampled in a sequencing experiment. (a–d) Possible configurations of germ and sampled somatic cellular lineages at the early post-zygotic stage: (a) any cell ancestral to sampled somatic and germ cells is ancestral to all such cells; (b) cells may be ancestral to all germ cells but only some sampled somatic cells; (c) cells may be ancestral to all sampled somatic cells but only some germ cells; (d) cells may be ancestral to some germ cells and some sampled somatic cells (meaning that some germ cells may be more closely related to some somatic cells than to other germ cells, and vice versa).
Abstract
Genome sequencing studies of de novo mutations in humans have revealed surprising incongruities in our understanding of human germline mutation. In particular, the mutation rate observed in modern humans is substantially lower than that estimated from calibration against the fossil record, and the paternal age effect in mutations transmitted to offspring is much weaker than expected from our long-standing model of spermatogenesis. I consider possible explanations for these discrepancies, including evolutionary changes in life-history parameters such as generation time and the age of puberty, a possible contribution from undetected post-zygotic mutations early in embryo development, and changes in cellular mutation processes at different stages of the germline. I suggest a revised model of stem-cell state transitions during spermatogenesis, in which ‘dark’ gonial stem cells play a more active role than hitherto envisaged, with a long cycle time undetected in experimental observations. More generally, I argue that the mutation rate and its evolution depend intimately on the structure of the germline in humans and other primates.
This article is part of the themed issue ‘Dating species divergences using rocks and clocks'.
FREE PDF GRATIS: Phil Trans R Soc B
Fechando a lacuna entre rochas e relógios usando evidência total de datação
Closing the gap between rocks and clocks using total-evidence dating
Fredrik Ronquist, Nicolas Lartillot, Matthew J. Phillips
Published 20 June 2016. DOI: 10.1098/rstb.2015.0136

Inferred phylogenetic affinities of fossils. Despite the plasticity in the morphological data and/or the imperfection of our models of morphological evolution, the placements of fossils suggested by total-evidence analyses largely agree with the views expressed by palaeontologists (see text for more detailed discussion). Results are shown for the model assuming rapid diversification, but fossil placements were similar for other informative priors on the diversification and fossil sampling processes.
Abstract
Total-evidence dating (TED) allows evolutionary biologists to incorporate a wide range of dating information into a unified statistical analysis. One might expect this to improve the agreement between rocks and clocks but this is not necessarily the case. We explore the reasons for such discordance using a mammalian dataset with rich molecular, morphological and fossil information. There is strong conflict in this dataset between morphology and molecules under standard stochastic models. This causes TED to push divergence events back in time when using inadequate models or vague priors, a phenomenon we term ‘deep root attraction’ (DRA). We identify several causes of DRA. Failure to account for diversified sampling results in dramatic DRA, but this can be addressed using existing techniques. Inadequate morphological models also appear to be a major contributor to DRA. The major reason seems to be that current models do not account for dependencies among morphological characters, causing distorted topology and branch length estimates. This is particularly problematic for huge morphological datasets, which may contain large numbers of correlated characters. Finally, diversification and fossil sampling priors that do not incorporate all the available background information can contribute to DRA, but these priors can also be used to compensate for DRA. Specifically, we show that DRA in the mammalian dataset can be addressed by introducing a modest extra penalty for ghost lineages that are unobserved in the fossil record, for instance by assuming rapid diversification, rare extinction or high fossil sampling rate; any of these assumptions produces highly congruent divergence time estimates with a minimal gap between rocks and clocks. Under these conditions, fossils have a stabilizing influence on divergence time estimates and significantly increase the precision of those estimates, which are generally close to the dates suggested by palaeontologists.
This article is part of the themed issue ‘Dating species divergences using rocks and clocks’.
FREE PDF GRATIS: Phil Trans R Soc B
Cálculo filogenético bayesiano de idades de fósseis
Bayesian phylogenetic estimation of fossil ages
Alexei J. Drummond, Tanja Stadler
Published 20 June 2016. DOI: 10.1098/rstb.2015.0129
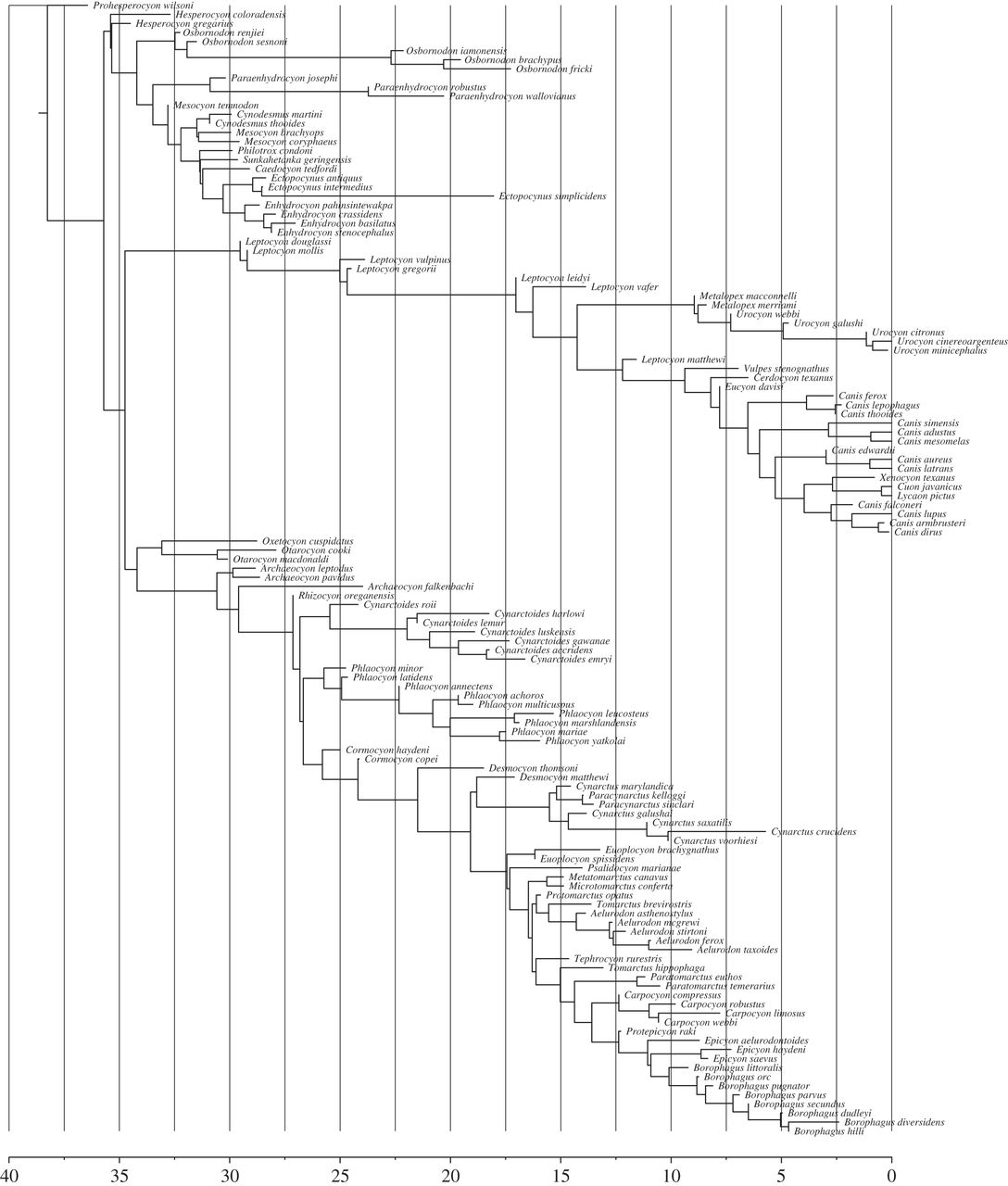
A sample from the posterior distribution of an analysis of the canid dataset, showing three main clades, one clade with extant representatives and two extinct clades (Hesperocyoninae and Borophaginae). The x-axis is in units of million years ago.
Abstract
Recent advances have allowed for both morphological fossil evidence and molecular sequences to be integrated into a single combined inference of divergence dates under the rule of Bayesian probability. In particular, the fossilized birth–death tree prior and the Lewis-Mk model of discrete morphological evolution allow for the estimation of both divergence times and phylogenetic relationships between fossil and extant taxa. We exploit this statistical framework to investigate the internal consistency of these models by producing phylogenetic estimates of the age of each fossil in turn, within two rich and well-characterized datasets of fossil and extant species (penguins and canids). We find that the estimation accuracy of fossil ages is generally high with credible intervals seldom excluding the true age and median relative error in the two datasets of 5.7% and 13.2%, respectively. The median relative standard error (RSD) was 9.2% and 7.2%, respectively, suggesting good precision, although with some outliers. In fact, in the two datasets we analyse, the phylogenetic estimate of fossil age is on average less than 2 Myr from the mid-point age of the geological strata from which it was excavated. The high level of internal consistency found in our analyses suggests that the Bayesian statistical model employed is an adequate fit for both the geological and morphological data, and provides evidence from real data that the framework used can accurately model the evolution of discrete morphological traits coded from fossil and extant taxa. We anticipate that this approach will have diverse applications beyond divergence time dating, including dating fossils that are temporally unconstrained, testing of the ‘morphological clock', and for uncovering potential model misspecification and/or data errors when controversial phylogenetic hypotheses are obtained based on combined divergence dating analyses.
This article is part of the themed issue ‘Dating species divergences using rocks and clocks’.
FREE PDF GRATIS: Phil Trans R Soc B
Um modelo de relógio misto livre
A mixed relaxed clock model
Nicolas Lartillot, Matthew J. Phillips, Fredrik Ronquist
Published 20 June 2016. DOI: 10.1098/rstb.2015.0132

Inferred divergence times under the mixed clock, using the tip-dating approach (posterior median, coloured bars: 95% credible intervals; red bars: Placentalia, as well as orders and super-orders indicated to the right).
Abstract
Over recent years, several alternative relaxed clock models have been proposed in the context of Bayesian dating. These models fall in two distinct categories: uncorrelated and autocorrelated across branches. The choice between these two classes of relaxed clocks is still an open question. More fundamentally, the true process of rate variation may have both long-term trends and short-term fluctuations, suggesting that more sophisticated clock models unfolding over multiple time scales should ultimately be developed. Here, a mixed relaxed clock model is introduced, which can be mechanistically interpreted as a rate variation process undergoing short-term fluctuations on the top of Brownian long-term trends. Statistically, this mixed clock represents an alternative solution to the problem of choosing between autocorrelated and uncorrelated relaxed clocks, by proposing instead to combine their respective merits. Fitting this model on a dataset of 105 placental mammals, using both node-dating and tip-dating approaches, suggests that the two pure clocks, Brownian and white noise, are rejected in favour of a mixed model with approximately equal contributions for its uncorrelated and autocorrelated components. The tip-dating analysis is particularly sensitive to the choice of the relaxed clock model. In this context, the classical pure Brownian relaxed clock appears to be overly rigid, leading to biases in divergence time estimation. By contrast, the use of a mixed clock leads to more recent and more reasonable estimates for the crown ages of placental orders and superorders. Altogether, the mixed clock introduced here represents a first step towards empirically more adequate models of the patterns of rate variation across phylogenetic trees.
This article is part of the themed issue ‘Dating species divergences using rocks and clocks’.
FREE PDF GRATIS: Phil Trans R Soc B
Uma exploração molecular paleobiológica da terrestrialização dos artrópodes
A molecular palaeobiological exploration of arthropod terrestrialization
Jesus Lozano-Fernandez, Robert Carton, Alastair R. Tanner, Mark N. Puttick, Mark Blaxter, Jakob Vinther, Jørgen Olesen, Gonzalo Giribet, Gregory D. Edgecombe, Davide Pisani
Published 20 June 2016. DOI: 10.1098/rstb.2015.0133

Bayesian phylogeny of Panarthropoda. This tree was obtained under the CAT − GTR + G model. All nodes but one had a posterior probability of 1. bpcomp maxdiff = 0; minimum effective size = 55; maximum rel_diff = 0.2. Most silhouettes from organisms are from Phylopic (phylopic.org/).
Abstract
Understanding animal terrestrialization, the process through which animals colonized the land, is crucial to clarify extant biodiversity and biological adaptation. Arthropoda (insects, spiders, centipedes and their allies) represent the largest majority of terrestrial biodiversity. Here we implemented a molecular palaeobiological approach, merging molecular and fossil evidence, to elucidate the deepest history of the terrestrial arthropods. We focused on the three independent, Palaeozoic arthropod terrestrialization events (those of Myriapoda, Hexapoda and Arachnida) and showed that a marine route to the colonization of land is the most likely scenario. Molecular clock analyses confirmed an origin for the three terrestrial lineages bracketed between the Cambrian and the Silurian. While molecular divergence times for Arachnida are consistent with the fossil record, Myriapoda are inferred to have colonized land earlier, substantially predating trace or body fossil evidence. An estimated origin of myriapods by the Early Cambrian precedes the appearance of embryophytes and perhaps even terrestrial fungi, raising the possibility that terrestrialization had independent origins in crown-group myriapod lineages, consistent with morphological arguments for convergence in tracheal systems.
This article is part of the themed issue ‘Dating species divergences using rocks and clocks’.
FREE PDF GRATIS: Phil Trans R Soc B
Blocos tectônicos e relógios moleculares
Tectonic blocks and molecular clocks
Kenneth De Baets, Alexandre Antonelli, Philip C. J. Donoghue
Published 20 June 2016. DOI: 10.1098/rstb.2016.0098

Possible relationship between divergences of terrestrial groups with different dispersal abilities and the age constraints from the break-up of continents and formation of oceans. Continent reconstructions are simplified after Stanley & Luczaj [37].
Abstract
Evolutionary timescales have mainly used fossils for calibrating molecular clocks, though fossils only really provide minimum clade age constraints. In their place, phylogenetic trees can be calibrated by precisely dated geological events that have shaped biogeography. However, tectonic episodes are protracted, their role in vicariance is rarely justified, the biogeography of living clades and their antecedents may differ, and the impact of such events is contingent on ecology. Biogeographic calibrations are no panacea for the shortcomings of fossil calibrations, but their associated uncertainties can be accommodated. We provide examples of how biogeographic calibrations based on geological data can be established for the fragmentation of the Pangaean supercontinent: (i) for the uplift of the Isthmus of Panama, (ii) the separation of New Zealand from Gondwana, and (iii) for the opening of the Atlantic Ocean. Biogeographic and fossil calibrations are complementary, not competing, approaches to constraining molecular clock analyses, providing alternative constraints on the age of clades that are vital to avoiding circularity in investigating the role of biogeographic mechanisms in shaping modern biodiversity.
This article is part of the themed issue ‘Dating species divergences using rocks and clocks’.
FREE PDF GRATIS: Phil Trans R Soc B
A evolução dos métodos para estabelecer períodos evolutivos
The evolution of methods for establishing evolutionary timescales
Philip C. J. Donoghue, Ziheng Yang
Published 20 June 2016. DOI: 10.1098/rstb.2016.0020

Tip-calibration relies upon a molecular sequence alignment from living species, a morphological character set for living and fossil species, and a prior topology (a); total-evidence dating co-estimates topology and timescale. Branch lengths are estimated in a Bayesian MCMC approach based on both data types for living lineages and based on morphological data alone for the extinct lineages; these are calibrated to time based on the age of the fossil species (b). The divergence time estimates and inferred rates of molecular and morphological evolution are based on a consensus of the MCMC analysis (c)
Abstract
The fossil record is well known to be incomplete. Read literally, it provides a distorted view of the history of species divergence and extinction, because different species have different propensities to fossilize, the amount of rock fluctuates over geological timescales, as does the nature of the environments that it preserves. Even so, patterns in the fossil evidence allow us to assess the incompleteness of the fossil record. While the molecular clock can be used to extend the time estimates from fossil species to lineages not represented in the fossil record, fossils are the only source of information concerning absolute (geological) times in molecular dating analysis. We review different ways of incorporating fossil evidence in modern clock dating analyses, including node-calibrations where lineage divergence times are constrained using probability densities and tip-calibrations where fossil species at the tips of the tree are assigned dates from dated rock strata. While node-calibrations are often constructed by a crude assessment of the fossil evidence and thus involves arbitrariness, tip-calibrations may be too sensitive to the prior on divergence times or the branching process and influenced unduly affected by well-known problems of morphological character evolution, such as environmental influence on morphological phenotypes, correlation among traits, and convergent evolution in disparate species. We discuss the utility of time information from fossils in phylogeny estimation and the search for ancestors in the fossil record.
This article is part of the themed issue ‘Dating species divergences using rocks and clocks’.
FREE PDF GRATIS: Phil Trans R Soc B
A Royal Society vai discutir novas tendências em biologia evolucionária: 7-9 de novembro de 2016
terça-feira, junho 28, 2016
New trends in evolutionary biology: biological, philosophical and social science perspectives
Scientific meeting
Starts: November 07 2016 09:00 Add to calendar
Ends: November 09 2016 17:00 Add to calendar

Location
The Royal Society, London, 6-9 Carlton House Terrace, London, SW1Y 5AG
Overview
Scientific discussion meeting organised in partnership with the British Academy by Professor Denis Noble CBE FMedSci FRS, Professor Nancy Cartwright, Professor Sir Patrick Bateson FRS, Professor John Dupré and Professor Kevin Laland.
Developments in evolutionary biology and adjacent fields have produced calls for revision of the standard theory of evolution, although the issues involved remain hotly contested. This meeting will present these developments and arguments in a form that will encourage cross-disciplinary discussion and, in particular, involve the humanities and social sciences in order to provide further analytical perspectives and explore the social and philosophical implications.
The schedule of talks, and biographies of the organisers and speakers are available below. Alternatively you can download the draft programme (PDF). Speaker abstracts will be available closer to the meeting date. Recorded audio of the talks will be available on this page after the event has taken place.
Attending this event
This event is intended for researchers in relevant fields and is free to attend, but advance registration is essential. An optional lunch can be purchased at the time of registration and is priced at £20 per day.
Enquiries: Contact the events team.
+++++
NOTA DESTE BLOGGER:
Em 1998 este blogger anunciou que uma iminente e eminente mudança paradigmática em biologia evolucionária estava por acontecer em razão das muitas anomalias e dificuldades que a teoria da evolução de Darwin - na sua versão Síntese Evolutiva Moderna enfrentava no contexto de justificação teórica desde os anos 1980s. Foi Stephen Jay Gould quem afirmou ser a teoria neodarwinista apenas ortodoxia nos livros didáticos, mas não entre os que praticavam ciência normal e a discutiam intramuros e em artigos científicos.
Em 1998 este blogger anunciou que uma iminente e eminente mudança paradigmática em biologia evolucionária estava por acontecer em razão das muitas anomalias e dificuldades que a teoria da evolução de Darwin - na sua versão Síntese Evolutiva Moderna enfrentava no contexto de justificação teórica desde os anos 1980s. Foi Stephen Jay Gould quem afirmou ser a teoria neodarwinista apenas ortodoxia nos livros didáticos, mas não entre os que praticavam ciência normal e a discutiam intramuros e em artigos científicos.
Apesar de ter me apoiado em um dos gigantes daquele paradigma evolutivo, este blogger foi vilipendiado por muitos cientistas da Nomenklatura científica e pela Galera dos meninos e meninas de Darwin. Sempre ressaltei aqui - eu fico muito lisonjeado quando sou vindicado por evolucionistas honestos. Imagine agora ser vindicado por um encontro de cientistas na The Royal Society de Londres, Inglaterra. O coração bate rucututuco, e eu, não sei por que, rio daqueles que riram e zombaram do então simples professor de ensino médio, hoje mestre em História da Ciência pela PUC-São Paulo.
O bom é que a ciência e a mentira não podem andar de mãos dadas, pois a ciência é a busca pela verdade científica dos fatos encontrados na realidade. A mudança paradigmática já se deu em agosto de 2015 com o lançamento da nova teoria geral da evolução - a SÍNTESE EVOLUTIVA AMPLIADA/ESTENDIDA, que relegou a seleção natural a segundo plano e incorporou teses neolamarckistas. Mas o que significa tudo isso para o fato, Fato, FATO da evolução não foi contado aos alunos do ensino médio e superior, e nem é debatido em salas de aulas. Quanta bobagem de patrulhamento de conhecimentos que estão ao alcance de uma tecla de computador plugado na internet!
Parabéns, The Royal Society, pela iniciativa de demonstrar que Darwin não é corroborado no contexto de justificação teórica e algo precisa ser feito a respeito.
Darwin kaput desde 1859!!!
O bom é que a ciência e a mentira não podem andar de mãos dadas, pois a ciência é a busca pela verdade científica dos fatos encontrados na realidade. A mudança paradigmática já se deu em agosto de 2015 com o lançamento da nova teoria geral da evolução - a SÍNTESE EVOLUTIVA AMPLIADA/ESTENDIDA, que relegou a seleção natural a segundo plano e incorporou teses neolamarckistas. Mas o que significa tudo isso para o fato, Fato, FATO da evolução não foi contado aos alunos do ensino médio e superior, e nem é debatido em salas de aulas. Quanta bobagem de patrulhamento de conhecimentos que estão ao alcance de uma tecla de computador plugado na internet!
Parabéns, The Royal Society, pela iniciativa de demonstrar que Darwin não é corroborado no contexto de justificação teórica e algo precisa ser feito a respeito.
Darwin kaput desde 1859!!!
Debate da Síntese Evolutiva Ampliada/Estendida: onde a ciência se encontra com a filosofia, mas os alunos não ficam sabendo!
segunda-feira, junho 27, 2016
The Extended (Evolutionary) Synthesis Debate: Where Science Meets Philosophy
Massimo Pigliucci and Leonard Finkelman
- Author Affiliations
Massimo Pigliucci (massimo@platofootnote.org) and Leonard Finkelman are affiliated with the Philosophy Program in the City University of New York's Graduate Center.

Source/Fonte: PNAS Bernardi 2007
Abstract
Recent debates between proponents of the modern evolutionary synthesis (the standard model in evolutionary biology) and those of a possible extended synthesis are a good example of the fascinating tangle among empirical, theoretical, and conceptual or philosophical matters that is the practice of evolutionary biology. In this essay, we briefly discuss two case studies from this debate, highlighting the relevance of philosophical thinking to evolutionary biologists in the hope of spurring further constructive cross-pollination between the two fields.
Key words: modern synthesis extended synthesis philosophy of science epigenetic inheritance levels of selection
FREE PDF GRATIS: BioScience
Insetos já "usavam" camuflagem há 100 milhões de anos atrás!
Debris-carrying camouflage among diverse lineages of Cretaceous insects
Bo Wang1,2,3,*, Fangyuan Xia4, Michael S. Engel5, Vincent Perrichot6, Gongle Shi1, Haichun Zhang1, Jun Chen1,7, Edmund A. Jarzembowski1,8, Torsten Wappler2 and Jes Rust2
- Author Affiliations
1State Key Laboratory of Palaeobiology and Stratigraphy, Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing 210008, China.
2Steinmann Institute, University of Bonn, 53115 Bonn, Germany.
3Key Laboratory of Zoological Systematics and Evolution, Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China.
4Nanjiao Bieshu 394, Shanghai 201108, China.
5Division of Entomology, Natural History Museum and Department of Ecology and Evolutionary Biology, University of Kansas, Lawrence, KS 66045, USA.
6CNRS UMR 6118 Géosciences and OSUR, Université de Rennes 1, 35042 Rennes, France.
7Institute of Geology and Paleontology, Linyi University, Linyi 276000, China.
8Department of Earth Sciences, Natural History Museum, London SW7 5BD, UK.
↵*Corresponding author. Email: bowang@nigpas.ac.cn
Science Advances 24 Jun 2016: Vol. 2, no. 6, e1501918

Fig. 1
Chrysopoid larvae from Mid-Cretaceous Burmese amber.
(A) Morphotype CI, BA12018. An, antenna; Ba, barklouse; He, head; Ja, jaw; Ps, psyllid; Le, leg; Lt, long tubular tubercle; St, short tubular tubercle. Note the two arthropod exoskeletons (psyllid and barklouse) attached to tubular tubercles. (B) Morphotype CII, L14002, naked. (C) Morphotype CIII, BA12019, naked. (D) Morphotype CII, NIGP164061, camouflaged. (E) Morphotype CII, NIGP164054, camouflaged. Scale bars, 2 mm (A) and 1 mm (B to E).
Abstract
Insects have evolved diverse methods of camouflage that have played an important role in their evolutionary success. Debris-carrying, a behavior of actively harvesting and carrying exogenous materials, is among the most fascinating and complex behaviors because it requires not only an ability to recognize, collect, and carry materials but also evolutionary adaptations in related morphological characteristics. However, the fossil record of such behavior is extremely scarce, and only a single Mesozoic example from Spanish amber has been recorded; therefore, little is known about the early evolution of this complicated behavior and its underlying anatomy. We report a diverse insect assemblage of exceptionally preserved debris carriers from Cretaceous Burmese, French, and Lebanese ambers, including the earliest known chrysopoid larvae (green lacewings), myrmeleontoid larvae (split-footed lacewings and owlflies), and reduviids (assassin bugs). These ancient insects used a variety of debris material, including insect exoskeletons, sand grains, soil dust, leaf trichomes of gleicheniacean ferns, wood fibers, and other vegetal debris. They convergently evolved their debris-carrying behavior through multiple pathways, which expressed a high degree of evolutionary plasticity. We demonstrate that the behavioral repertoire, which is associated with considerable morphological adaptations, was already widespread among insects by at least the Mid-Cretaceous. Together with the previously known Spanish specimen, these fossils are the oldest direct evidence of camouflaging behavior in the fossil record. Our findings provide a novel insight into early evolution of camouflage in insects and ancient ecological associations among plants and insects.
Keywords Cretaceous paleoecology paleoentomology paleobotany camouflage
Copyright © 2016, The Authors
This is an open-access article distributed under the terms of the Creative Commons Attribution-NonCommercial license, which permits use, distribution, and reproduction in any medium, so long as the resultant use is not for commercial advantage and provided the original work is properly cited.
FREE PDF GRATIS: Science Advances Sup Info
Dorvillé, em vez de publicar sobre a tensão ciência-religião, que tal abordar a falência epistêmica da teoria da evolução de Darwin no contexto de justificação teórica?
domingo, junho 26, 2016
Luís Fernando Marques Dorvillé na sua tese de doutorado Religião, escola e ciência: Conflitos e tensões nas visões de mundo de alunos de uma licenciatura em ciências biológicas (2010), e no seu artigo Criacionismo: transformações históricas e implicações para o ensino de ciências e biologia (2016), onde os termos identificando a teoria do Design Inteligente (TDI) e a Sociedade Brasileira de Design Inteligente foram inapropriadamente usados: desenho inteligente.
Na sua tese de doutorado na Universidade Federal Fluminense, o termo inadequado "desenho inteligente" foi usado 50 vezes. Na página 120, ele usou corretamente o termo "design inteligente" identificando este blogger como então coordenador do NBDI (Núcleo Brasileiro de Design Inteligente) que, posteriormente, deu origem à SBDI (Sociedade Brasileira do Design Inteligente). Além dessa menção, mais três foram devidamente mantidas. Repare a data mencionada no texto dele: 1998. Desde lá a nomenclatura acolhida é "design inteligente" e não desenho inteligente. Razão? Desconhecemos nas faculdades públicas e privada um curso de Design ser rotulado de Desenho. Pode indicar-me uma? Nem perca seu tempo, Dorvillé - não existe uma assim faculdade de Design assim designada!
No seu artigo recente - Criacionismo: transformações históricas e implicações para o ensino de ciências e biologia, o termo "desenho inteligente" foi usado 11 vezes, e na página 486, a SBDI foi erroneamente mencionada como Sociedade Brasileira do Desenho Inteligente, quando o correto é Sociedade Brasileira do Design Inteligente. Ao mencionar a data - 2014, ele teve acesso a algum documento sobre o evento que a Grande Mídia reportou corretamente. Não seguir a designação correta, mostra uma indisposição intencional em descaracterizar um movimento científico.
Tanto na sua tese como no artigo percebe-se uma nítida aversão pelos cristãos evangélicos, quando em nosso movimento nós temos ateus, agnósticos, sem religião, e cristãos, e isso não aparece destacado em sua obra. Mais sofrível ainda é encontrar na bibliografia de sua tese de doutorado somente, NOTA BENE, somente um livro de autor ligado ao Design Inteligente: Michael Behe. Há outros autores, e o mais expoente - William Dembski, filósofo e matemático, ficou de fora. Numa tese de doutorado, é de suma importância "ouvir o outro lado" e reportar suas teses como expostas pelos seus teóricos e defensores, e daí concordar, discordar e criticar suas afirmações. Não se vê isso na tese de Dorvillé, que se apoiou exclusivamente em literatura crítica à TDI - quer dizer, informação de segunda mão, que não cabe numa tese de doutorado. E o orientador nem se deu conta disso? E a banca que aprovou a tese nem percebeu uma lacuna bibliográfica dessa monta?
Dorvillé, a TDI não é criacionismo, e segundo Ronald Numbers, um dos autores amplamente usado por ele, e crítico do DI, afirmou que o termo “criacionismo” é inexato quando se refere ao movimento do DI. Então por que a insistência em nos rotular assim? Segundo Numbers, é porque os críticos e oponentes pensam que tais afirmações são a forma mais fácil em desacreditar o DI. A TDI é uma teoria científica minimalista que afirma existir sinais de inteligência na natureza que são empiricamente detectados, tais como complexidade irredutível de sistemas biológicos e informação complexa especificada, e isso derivado do que foi encontrado na natureza, e não em relatos de criação de livros considerados sagrados pelos crentes de subjetividades religiosas.
Fique despreocupado, Dorvillé, nós do movimento do Design Inteligente, somos contra o ensino da TDI nas escolas e universidades públicas e privadas, não somente no Brasil, mas no mundo inteiro, enquanto ela não for amplamente debatida e aceita pela comunidade científica. Vide abaixo o nosso manifesto a respeito. E não dependemos da falência epistemológica da teoria da evolução de Darwin para que a nossa teoria seja cientificamente estabelecida.
Enquanto isso não se dá, expomos sim a falência epistemológica das teorias da evolução no contexto de justificação teórica porque Dorvillé e os demais que praticam ciência normal não ousam fazê-lo, e isso é compreensível: o que vale é o paradigma, as evidências contrárias que se danem! E há uma carreira a preservar, por que correr o risco de desafiar a Nomenklatura científica, não é Dorvillé?
+++++
MANIFESTO PÚBLICO DA SOCIEDADE BRASILEIRA DO DESIGN INTELIGENTE -TDI BRASIL- SOBRE O ENSINO DA TEORIA DA EVOLUÇÃO E DA TEORIA DO DESIGN INTELIGENTE NAS ESCOLAS E UNIVERSIDADES PÚBLICAS E PRIVADAS
"A TDI-BRASIL declara, como sua política educacional, não ser favorável, na atual conjuntura acadêmica, ao ensino da Teoria do Design Inteligente (TDI) nas escolas e universidades brasileiras públicas e privadas, como também nas confessionais.
Nossa posição se fundamenta na opinião atual da Academia, que ainda não acata em sua maioria a TDI e o seu ensino, posição essa que nós da TDI BRASIL, como acadêmicos, devemos acatar.
Outro fundamento de nossa posição contrária ao ensino da TDI nas escolas é a não existência, no quadro educacional atual, de professores capacitados para corretamente ensinar os postulados da TDI.
Entendemos, porém, que os alunos têm o direito constitucional de ser informados que há uma disputa já instalada na academia entre a teoria da evolução (TE) e a TDI quanto à melhor inferência científica sobre nossas origens. Inclusive há outras correntes acadêmicas, além da TDI, que hoje questionam a validade da TE oferecendo uma terceira via.
Quanto ao ensino da TE, a TDI BRASIL defende que este ensino seja feito, porém, de uma forma honesta e imparcial, tanto nos livros didáticos quanto na exposição dos professores em salas de aula. Defendemos que sejam eliminados exemplos fraudulentos ou equivocados atualmente presentes em livros didáticos, e que sejam expostas as deficiências graves que a TE apresenta, e que se agravam a cada dia frente às descobertas científicas mais recentes - o que hoje não ocorre.
Quanto ao criacionismo, na sua versão religiosa e filosófica, por causa de seus pressupostos filosóficos e teológicos, entendemos que deva ser ensinado e discutido, junto com as evidências científicas que porventura o corroborem, em aulas de filosofia e teologia, dando a estas disciplinas o seu devido valor no debate sobre as nossas origens."
A percepção da teoria do Design Inteligente cresce no Brasil e preocupa a Nomenklatura científica
sexta-feira, junho 24, 2016
A Percepção Sobre a Hipótese do Design Inteligente no Brasil (Minas Gerais)
The perception of the hypothesis of Intelligent Design in Brazil (Minas Gerais)
Heslley Machado Silva 1, Graça S. Carvalho 2, Paloma Rodrigues da Silva 3, Daiana Evilin Gibram 4
1 Centro Universitário de Formiga, Minas Gerais, Brasil. Universidade de Itaúna, Minas Gerais , Brasil
² Instituto de Investigação da Universidade do Minho, Portugal.
³ Universidade Estadual Paulista, São Paulo, Brasil.
4 Centro Universitário de Formiga, Minas Gerais, Brasil.

Resumo
Introdução: Este trabalho é um recorte de uma pesquisa mais realizada no Brasil, que fez um paralelo entre um relatório recentemente publicado no Reino Unido, intitulado Rescuing Darwin, que trata de como a população britânica percebe a questão da evolução biológica e temas relacionados.
Objetivo: O recorte apresenta e analisa os resultados referentes ao tema do design inteligente. Em ambos os países a maioria dos entrevistados acredita nessa hipótese [sic], com notável penetração no Brasil. A hipótese tem ampla aceitação nos dois gêneros, com leve superioridade no sexo feminino.
Resultados: Entre as principais religiões brasileiras houve grande aceitação entre os evangélicos, seguidos dos católicos e também os espíritas. Analisou-se também a influência do nível de escolaridade. Não há mudança significativa quando se aumenta o grau de escolaridade, sendo que em todos a aceitação da hipótese do design inteligente é alta.
Conclusão: A partir dessa percepção, urge discutir como o conhecimento científico é apresentado à população, o grau de compreensão dessa visão e como a população a vê diante da evolução darwinista, tão aceita cientificamente. Por fim, analisa-se como esse quadro é preocupante diante de ações que ocorrem no mundo e no Brasil, que visam alterar o ensino de evolução biológica, substituindo-o por uma abordagem dogmática, com preceitos religiosos que estão dissociados da metodologia científica. [???]
Palavras-chave: Educação. Evolução. Design Inteligente.
Autor correspondente:
Heslley Machado Silva
E-mail: heslley@uniformg.edu.br
Recebido em: 23/10/2015 Revisado em: 04/05/2016 Aceito em: 06/05/2016
Publicado em: 15/06/2016
+++++
NOTA CAUSTICANTE DESTE BLOGGER:
Heslley Machado Silva é um pesquisador com uma agenda, não é um cientista sério que segue as evidências aonde elas forem dar, e sequer tem conhecimento mínimo do que seja a teoria (que ele minimiza como hipótese) do Design Inteligente e sua política quanto ao seu ensino nas escolas e universidades públicas e privadas. A foto acima corrobora a conclusão de Heslley: a penetração da TDI no Brasil é notável. Não somente isso, é irreversível.
O manifesto lançado naquele evento em Campinas, SP, demonstra quão em descompasso com a verdade ele está em relação aos mais de 350 professores e pesquisadores, alguns em universidades brasileiras públicas e privadas de renome, e a nossa política de ensino da TDI, nada parecido com o falso alarme dado por Heslley de ser um quadro preocupante diante de ações que ocorrem no mundo e no Brasil, que visam alterar o ensino de evolução biológica, substituindo-o por uma abordagem dogmática, com preceitos religiosos que estão dissociados da metodologia científica:
Heslley Machado Silva é um pesquisador com uma agenda, não é um cientista sério que segue as evidências aonde elas forem dar, e sequer tem conhecimento mínimo do que seja a teoria (que ele minimiza como hipótese) do Design Inteligente e sua política quanto ao seu ensino nas escolas e universidades públicas e privadas. A foto acima corrobora a conclusão de Heslley: a penetração da TDI no Brasil é notável. Não somente isso, é irreversível.
O manifesto lançado naquele evento em Campinas, SP, demonstra quão em descompasso com a verdade ele está em relação aos mais de 350 professores e pesquisadores, alguns em universidades brasileiras públicas e privadas de renome, e a nossa política de ensino da TDI, nada parecido com o falso alarme dado por Heslley de ser um quadro preocupante diante de ações que ocorrem no mundo e no Brasil, que visam alterar o ensino de evolução biológica, substituindo-o por uma abordagem dogmática, com preceitos religiosos que estão dissociados da metodologia científica:
Manifesto público da Sociedade Brasileira do Design Inteligente – TDI-Brasil – sobre o ensino da Teoria da Evolução e da Teoria do Design Inteligente nas escolas e universidades públicas e privadas
"A TDI-Brasil declara, como sua política educacional, não ser favorável, na atual conjuntura acadêmica, ao ensino da Teoria do Design Inteligente (TDI) nas escolas e universidades brasileiras públicas e privadas, como também nas confessionais.
Nossa posição se fundamenta na opinião atual da Academia, que ainda não acata em sua maioria a TDI e o seu ensino, posição essa que nós da TDI-Brasil, como acadêmicos, devemos acatar.
Outro fundamento de nossa posição contrária ao ensino da TDI nas escolas é a não existência, no quadro educacional atual, de professores capacitados para corretamente ensinar os postulados da TDI.
Entendemos, porém, que os alunos têm o direito constitucional de ser informados que há uma disputa já instalada na academia entre a teoria da evolução (TE) e a TDI quanto à melhor inferência científica sobre nossas origens. Inclusive há outras correntes acadêmicas, além da TDI, que hoje questionam a validade da TE oferecendo uma terceira via.
Quanto ao ensino da TE, a TDI-Brasil defende que este ensino seja feito, porém, de uma forma honesta e imparcial, tanto nos livros didáticos quanto na exposição dos professores em salas de aula. Defendemos que sejam eliminados exemplos fraudulentos ou equivocados atualmente presentes em livros didáticos, e que sejam expostas as deficiências graves que a TE apresenta, e que se agravam a cada dia frente às descobertas científicas mais recentes – o que hoje não ocorre.
Quanto ao criacionismo, na sua versão religiosa e filosófica, por causa de seus pressupostos filosóficos e teológicos, entendemos que deva ser ensinado e discutido, junto com as evidências científicas que porventura o corroborem, em aulas de Filosofia e Teologia, dando a estas disciplinas o seu devido valor no debate sobre as nossas origens."
Heslley, estamos de olho nos seus artigos que desvirtuam o corpus epistêmico da teoria do Design Inteligente, e sua motivação ideológica espúria patente em querer demonizar uma teoria científica associando-a ao criacionismo.
Assinar:
Comentários (Atom)



