
Click here/Clique aqui: (157) Do You Love Me? - YouTube




Science journal editors shouldn’t contribute to politicizing science
By GENEVIEVE P. KANTER OCTOBER 23, 2020

When the editors of some of the world’s leading science journals agree on something, it is generally safe to assume that they are correct. So when prominent journals like Science, Nature, and the New England Journal of Medicine recently published editorials excoriating President Trump’s deadly bungling of the pandemic response and suppression of scientific activity, the editors accurately spotlighted the troubling deficiencies of the current administration.
But in advocating against or endorsing a presidential candidate, these editors made a grave error. In taking this extraordinary step, they made themselves vulnerable to charges of bias, overstepped their roles as science editors, and succumbed to the politicization of science that they and many other scientists find so alarming.
At first glance, these appear to be similar to run-of-the-mill newspaper endorsements. This analogy, however, is not quite right. At a newspaper, there is a wall between the news operation and the editorial office. It exists to prevent biases of the editorial staff from influencing news reporting. No such wall exists for science journals. The editors who write the editorials are the same ones who evaluate manuscripts and make the final decisions on whether to publish them.
There’s another problem: This political advocacy unnecessarily invites allusions to cronyism, echoing a less savory time when wealthy newspaper owners used their editorial pages to extol the merits of their political chums. Indeed, because of fears surrounding the appearance of undue influence and bias, many newspapers in recent years have abandoned political endorsements.
Read more here: STAT News






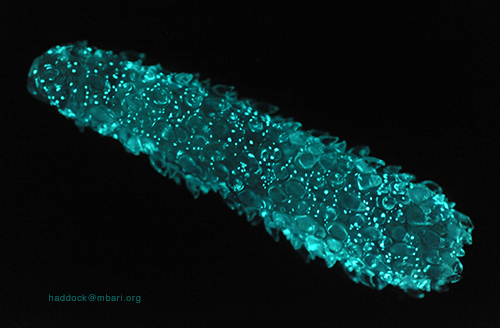
Pyrosomes, colonial salps, continue to be one of the most mysterious of bioluminescent organisms. Their glow can last 15 seconds or more, and it can be triggered by light, even cascading from one end of the colony to the other. The chemical origin remains unknown. It was thought to be bacterial, in part because of similar kinetics, but now appears to be intrinsic chemistry that lets this animal emit its impressive glow.

On the problem of biological form
Marta Linde-Medina
Theory in Biosciences volume 139, pages299–308(2020)
Abstract
Embryonic development, which inspired the first theories of biological form, was eventually excluded from the conceptual framework of the Modern Synthesis as irrelevant. A major question during the last decades has centred on understanding whether new advances in developmental biology are compatible with the standard view or whether they compel a new theory. Here, I argue that the answer to this question depends on which concept of morphogenesis is held. Morphogenesis can be conceived as (1) a chemically driven or (2) a mechanically driven process. According to the first option, genetic regulatory networks drive morphogenesis. According to the second, morphogenesis results from an invariant tendency of embryonic tissues to restore changes in mechanical stress. While chemically driven morphogenesis allows an extension of the standard view, mechanically driven morphogenesis would deeply transform it. Which of these hypotheses has wider explanatory power is unknown. At present, the problem of biological form remains unsolved.
Subscription or payment needed/Requer assinatura ou pagamento: Theory in Biosciences














{kind=link}